Главная
Цитоплазматическая мужская стерильность в баклажанах
Аннотация
Цитоплазматическая мужская стерильность (CMS) является полезной системой для производства гибридных семян у различных видов сельскохозяйственных культур. В баклажанах системы CMS были разработаны с использованием цитоплазмы шести диких видов Solanum путем повторных обратных скрещиваний. Эти системы CMS были классифицированы на два типа. Первый - это стерильность пыльника неделимого типа в системах CMS баклажана с цитоплазмами Solanum kurzii Brace & Prain, S. violaceum Ort. И S. virginianum L., в которых пыльник содержит нормальную пыльцу, но не раскрывается для высвобождения. , Второй тип - это бесплодие пыльцы не образующего типа в системах CMS баклажана с цитоплазмами S. aethiopicum Aculeatum Group, S. anguivi Lam. И S. grandifolium CV Morton, у которых пыльник мужской стерильной линии полностью лишен пыльцы. Оба типа системы бесплодия были охарактеризованы путем изучения фертильности пыльцы и семян. Кроме того, в системах CMS без образования пыльцы были обнаружены два независимых доминантных гена-восстановителя фертильности ( Rf ) и был разработан маркер секвенированной охарактеризованной амплифицированной области (SCAR), тесно связанный с этими генами.
Вступление
Баклажан ( Solanum melongena L.) является экономически важной овощной культурой семейства пасленовых , широко выращиваемой по всему миру. Эксплуатация гетерозиса стала потенциальным инструментом для улучшения баклажана ( Самбандам, 1962 ). В Японии, Нагаи и Кида (1926) первыми сообщили о количественных характеристиках гибридов баклажана и обнаружили, что гетерозис проявляется в общей урожайности, количестве плодов на растение, преждевременном цветении, преждевременной зрелости, высоте растения, количестве ветвей, количестве шипов на цветоножке и длина плода. Сообщалось о появлении замечательной гибридной силы в скрещиваниях на баклажанах в отношении массы семян, диаметра и высоты стебля, преждевременной продуктивности, большей энергии роста и более высокой урожайности ( Какидзаки, 1930 , 1931 ). Гото (1952) зарегистрировано заметное увеличение урожайности в поколении F1 серии скрещиваний между сортами японских баклажанов. Феномен гетерозиса или гибридной энергии в баклажанах хорошо известен сегодня, и селекционеры используют его для увеличения производства. Следовательно, большинство коммерческих сортов баклажанов в настоящее время являются межвариантными гибридами.
Мужское бесплодие включает неспособность растений производить функциональные пыльники, пыльцу или мужские гаметы, хотя потенциал для женского размножения остается. Цитоплазматическое мужское бесплодие (CMS) - это своего рода бесплодие у растений, которое вызывается специфическими ядерными и митохондриальными взаимодействиями. CMS является наследуемым по материнской линии признаком, который способствует производству гибридных семян многих культур и позволяет селекционерам использовать прирост урожайности, связанный с гибридной энергией. В настоящее время CMS была идентифицирована у более чем 150 видов растений ( Schnable and Wise, 1998 ). Хорошо документировано, что необычная открытая рамка считывания (ORF), состоящая из химерной структуры эндогенных митохондриальных генов в митохондриальном геноме CMS, ответственна за CMS ( Хансон и Бентолила, 2004 ). Во многих случаях было обнаружено, что мужская фертильность может быть восстановлена ядерным кодирующим геном ( Rf ) восстановителем фертильности, некоторые из которых были недавно выделены ( Бентолила и др., 2002 ). Ключевую роль в ядерно-генетическом контроле CMS играют Rf- гены, которые отвечают за восстановление мужской фертильности при взаимодействии с CMS-индуцирующей цитоплазмой. Эти гены, которые кодируют пентатрикопептидные повторяющиеся белки, влияют на экспрессию гена CMS посттранскрипционно. Это приводит к исчезновению белка CMS, по крайней мере, в мужских органах. Следовательно, системы CMS / Rf представляют ценность для изучения взаимодействий между ядерным и митохондриальным геномами.
Мужское бесплодие в баклажанах было описано в нескольких отчетах. Сообщалось о мужском бесплодии, вызванном рецессивными ядерными генами ( Чаухан, 1984 ; Жасмин, 1954 ; Наттол, 1963 ; Phatak and Jaworski, 1989 ; Phatak et al., 1991 ). CMS получен с использованием цитоплазмы дикого вида Solanum S. gilo Raddi ( Fang et al., 1985 ), S. kurzii Brace & Prain ( Хан и Иссики, 2009 ), S. violaceum Ort. ( Иссики и Каваджири, 2002 ), S. virginianum L. ( Хан и Иссики, 2008 ), S. aethiopicum Aculeatum Group ( Хан и Иссики, 2010 ), S. anguivi Lam. ( Хан и Иссики, 2011 ) и С. grandifolium CV Morton ( Хаснуннахар и др., 2012a ; Сайто и др., 2009а ) Сообщалось. В этом обзоре мы описываем цитоплазматическую мужскую стерильность баклажана, обнаруженную в нашем исследовании, уделяя особое внимание его развитию, характеристике, восстановлению фертильности и разработке маркера SCAR, связанного с генами Rf .
1. Разработка цитоплазматических систем мужской стерильности баклажана
1) Разработка пыльниковых неотделимых систем мужской стерильности
Были разработаны системы мужской стерильности баклажанов, не обладающие иммунитетом, с использованием цитоплазмы S. violaceum ( Иссики и Каваджири, 2002 ), S. virginianum ( Хан и Иссики, 2008 ) и С. Курзии ( Хан и Иссики, 2009 ). Межвидовые гибриды F1 между этими дикими видами Solanum и баклажанами были получены с использованием диких видов Solanum в качестве женских родителей и баклажана в качестве мужского. В результате повторных скрещиваний с баклажаном в качестве рекуррентного пыльцевого родителя были произведены обратные скрещивания. Все потомство обратного скрещивания выражало пыльность мужского бесплодия. Пыльник этих растений с мужской стерильностью (МС) не раскрывался для выделения пыльцы ( рисунок 1 ). Неисправность пыльников препятствовала выходу пыльцы из пыльников, хотя все потомство обратного скрещивания содержало нормальную пыльцу в своих пыльниках. Не пусковой характер пыльцы стал фиксированным в последующих поколениях обратного скрещивания. Известно, что в случае CMS степень мужской стерильности увеличивается с каждым последующим поколением обратного скрещивания ( МакВетти, 1997 ). Фертильность пыльцы оставалась ниже во всех потомствах обратного скрещивания этих трех систем CMS, чем в баклажанах ( рисунок 1 ). Линии MS показали нормальный мейоз и образование тетрады. Низкая фертильность пыльцы в этих линиях MS не может быть связана с мейотической трудностью. Характер пылеотделения и низкой фертильности пыльцы являются разновидностями ЦМС, вызванными дисгармонией между цитоплазмами диких видов Solanum и ядром S. melongena . Сообщалось, что некоторые системы CMS нестабильны при температуре и не пригодны для коммерческого использования для производства гибридных семян ( МакВетти, 1997 ); тем не менее, эти три линии MS показали устойчивую экспрессию пыльника нерешительного характера. Ни один ген-восстановитель фертильности этих трех систем CMS еще не обнаружен. Было обнаружено, что семенная фертильность линий МС очень высока при опылении баклажаном «Уттара» и почти равна плодовитости баклажана «Уттара» ( Таблица 1 ). Эти линии MS могут демонстрировать достаточную способность к семеноводству, когда они используются в качестве родительских семян для гибридного семеноводства. В баклажанах не было обнаружено отрицательного влияния цитоплазм S. kurzii , S. violaceum и S. virginianum на плодородие семян.
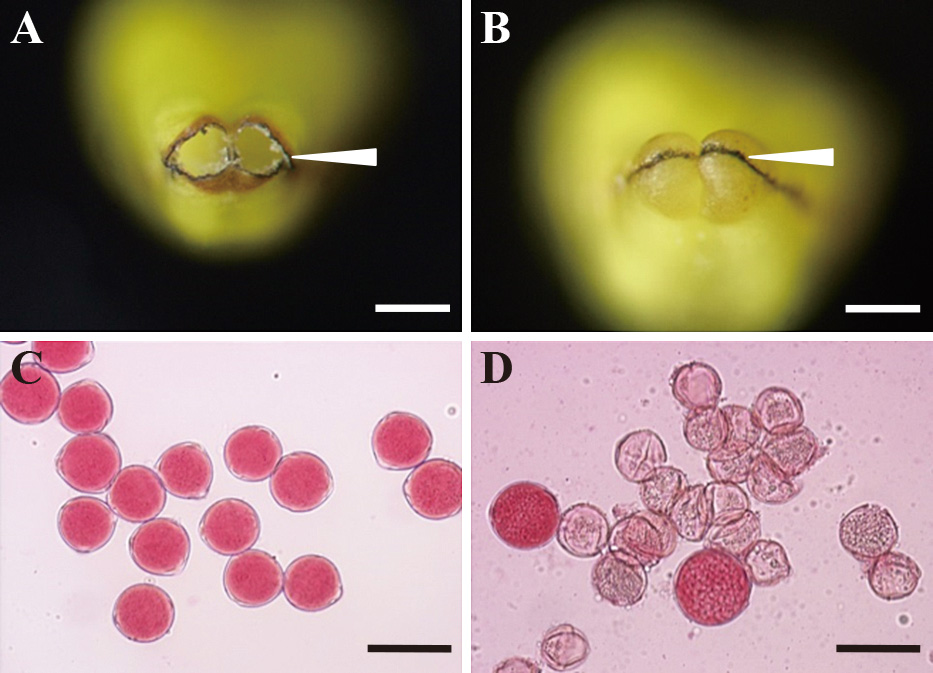
рисунок 1
Концы пыльника у цветущих (A, B) и окрашенных ацетокармином пыльцевых зерен (C, D) S. melongena 'Uttara' (слева) и растения BC4 с цитоплазмой S. virginianum (справа). Стрелками обозначена поровая часть пыльников. Шкала бар = 0,5 мм (A – B), 50 мкм (C – D).
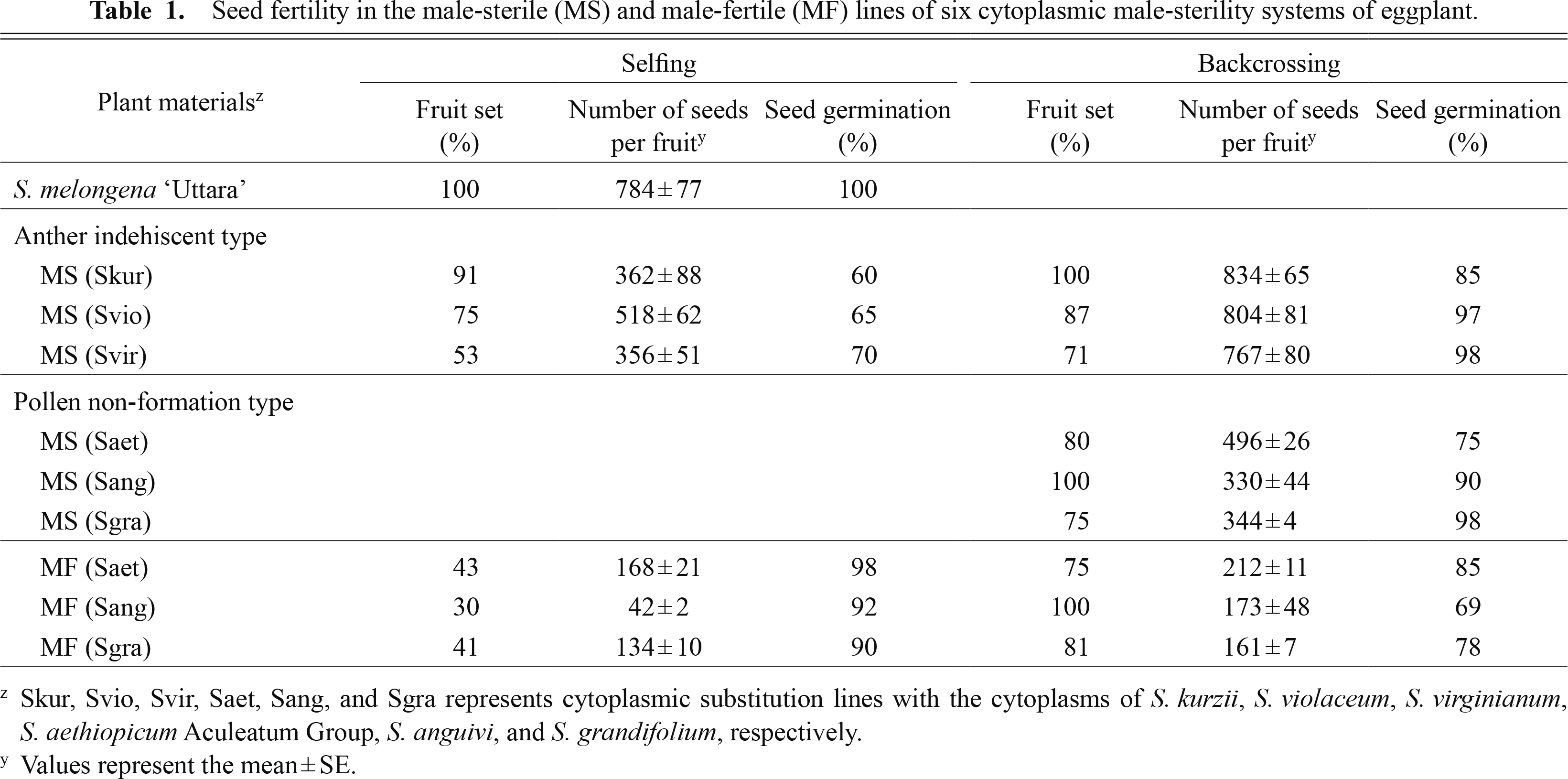
Таблица 1
Плодородие семян в линиях мужской стерильности (MS) и фертильности мужчин (MF) шести цитоплазматических систем мужской стерильности баклажана.
ПЦР-RFLP-анализ хлоропластной ДНК (кПДНК) и митохондриальной ДНК (мтДНК) показал материнское наследование в потомстве обратного скрещивания с цитоплазмами S. kurzii ( Хан и Иссики, 2009 ) и S. violaceum ( Иссики и Каваджири, 2002 ). ПЦР-RFLP анализ мтДНК показал материнское наследование в потомстве обратного скрещивания с цитоплазмой S. virginianum ( Хан и Иссики, 2008 ). Однако в случае анализа кПДНК методом ПЦР-RFLP исследуемое одиночное растение BC1 показало отцовские и новые рестрикционные паттерны, в дополнение к материнскому рестрикционному. Это указывало на возникновение бипарентального наследования кПДНК. Все последующее потомство обратного скрещивания показало одну и ту же модель рекомбинантной кДДНК растения BC1. Эти результаты позволяют предположить, что рекомбинантная кПДНК может быть стабильной и гармонизировать с ядерным геномом S. melongena .
2) Развитие систем мужской стерильности пыльцы без образования
Сообщалось, что системы CMS пыльцевого типа не образуют с использованием цитоплазмы S. aethiopicum Aculeatum Group ( Хан и Иссики, 2010 ), S. anguivi ( Хан и Иссики, 2011 ) и S. grandifolium ( Хаснуннахар и др., 2012a ; Сайто и др., 2009а ). Межвидовые гибриды F1 между дикими видами Solanum и баклажанами были получены с использованием диких видов Solanum в качестве женских родителей и баклажана в качестве мужского ( Рис. 2 ). Цитоплазма баклажана была заменена цитоплазмами диких видов Solanum путем повторного обратного скрещивания с использованием баклажана в качестве рецидивирующего пыльцевого родителя. Беккросс потомство S. aethiopicum Aculeatum Group, S. anguivi и S. grandifolium, индуцированных CMS, линии баклажана были разделены на MS без образования пыльцы и с формированием пыльцы мужского типа (MF) ( Таблица 2 ; Рис. 2 ). Мейоз не был обнаружен в линиях MS, тогда как он был обнаружен в материнских клетках пыльцы (PMC) линий MF. Пыльники линий MS были меньше по размеру, и пыльцевый мешок имел тенденцию быть черноватым с деформированной структурой и не содержал никакой пыльцы ( Рис. 3 ). Тем не менее, пыльник и пыльцевый мешок линий MF были нормальными, которые содержали значительную пыльцу ( Хан и Иссики, 2010 ). Линии MF имеют гены, восстанавливающие фертильность ( Rf ), которые способны противодействовать признаку мужской стерильности и восстанавливать фертильность пыльцы. Сообщалось, что два независимых доминантных гена-восстановителя фертильности контролируют образование пыльцы в этих трех системах CMS. У растений, содержащих ген Rf в одном или обоих из этих локусов, может образовываться пыльца, в то время как рецессивность обоих приводит к неспособности к образованию пыльцы. ПЦР-RFLP-анализ кпДНК и мтДНК показал материнское наследование в потомстве обратного скрещивания этих трех систем CMS. Это подтверждает, что метод повторного обратного скрещивания подходит для разработки линий замены цитоплазмы баклажана. Было обнаружено, что плодородие семян высоко в этих линиях CMS баклажана ( Таблица 1 ). В баклажанах не было обнаружено отрицательного влияния цитоплазм S. aethiopicum Aculeatum Group, S. anguivi и S. grandifolium на плодородие семян.
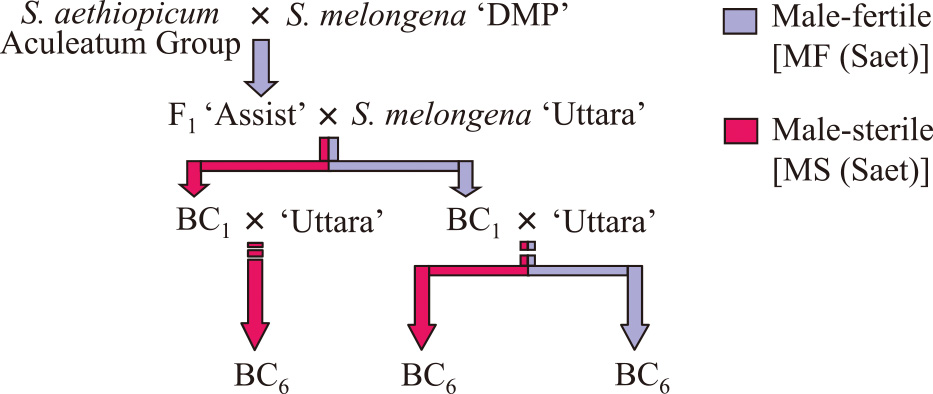
Рис. 2
Процедура замены цитоплазмы группы S. aethiopicum Aculeatum на группу S. melongena 'Uttara' непрерывным обратным скрещиванием.
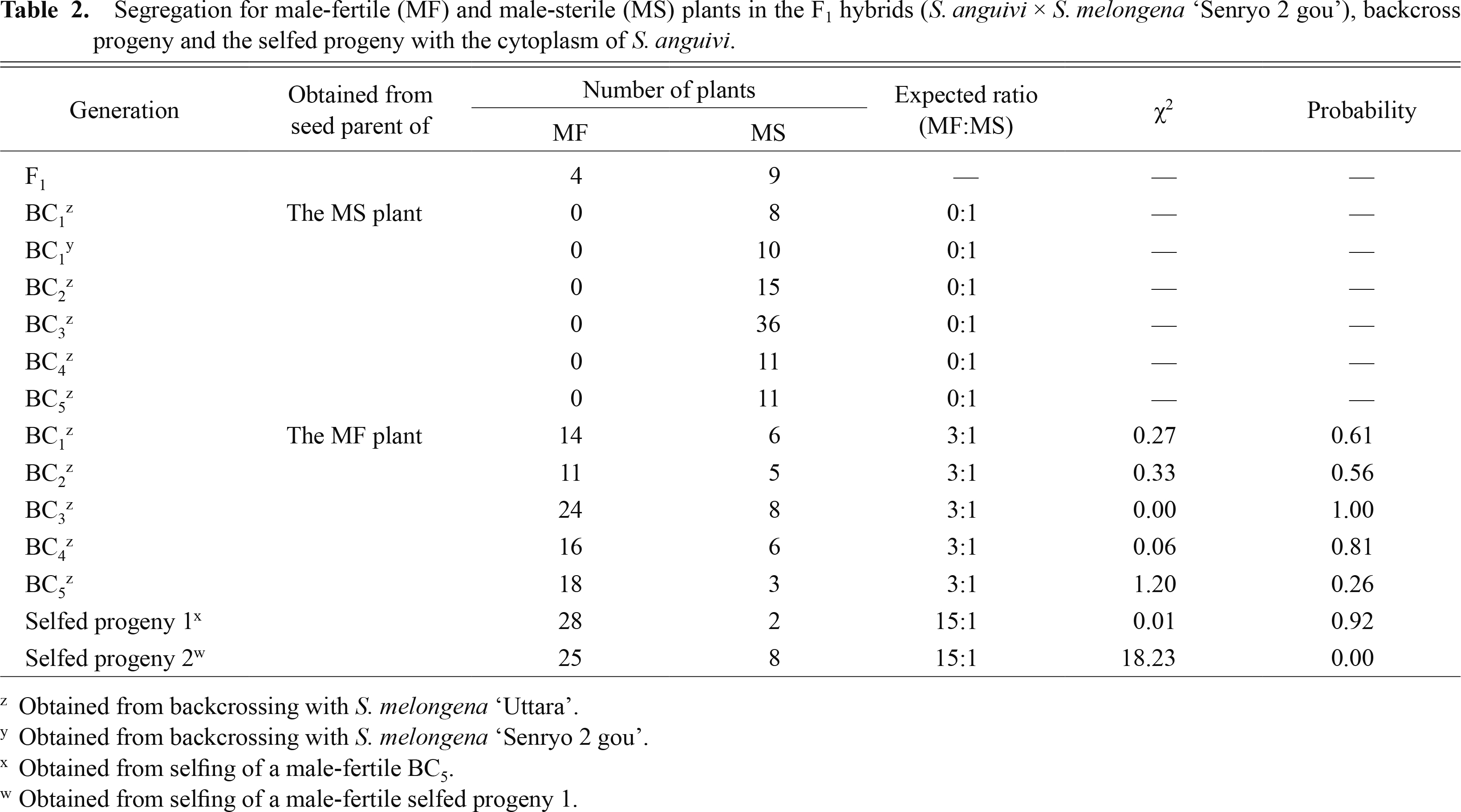
Таблица 2
Сегрегация для фертильных (MF) и стерильных (MS) растений у гибридов F1 ( S. anguivi × S. melongena 'Senryo 2 gou'), потомство обратного скрещивания и самоопыленное потомство с цитоплазмой S. anguivi .

Рис. 3
Цветы (A, B) и крупные поперечные разрезы пыльников (C, D) из мужских (слева) и мужских (стерильных) (справа) линий баклажана с цитоплазмой S. anguivi . Шкала бар = 10 мм (A – B), 0,5 мм (C – D).
2. Характеристика цитоплазматических систем мужской стерильности в баклажане
1) Характеристика пыльников нераздельных систем мужской стерильности
Характеристики пыльцы и плодородия семян были исследованы в системах пыльников мужского пола, не отличающихся друг от друга, разработанных с использованием цитоплазм S. kurzii , S. violaceum и S. virginianum, чтобы выяснить возможности размножения путем самоопыления и получения чистых линий посредством культура пыльников ( Хаснуннахар и др., 2012b ). Фертильность пыльцы оценивалась за 2 дня до введения, в день и через 2 дня после окрашивания по пыльцевым пятнам в ацетокармине и в растворе Люголя, а также по скорости прорастания пыльцы in vitro ( Рис. 4 а также 5 ). Окрашиваемость пыльцы ацетокармином и скорость прорастания пыльцы in vitro на всех трех стадиях созревания пыльцы были ниже в линиях MS, чем у S. melongena 'Uttara'. Окрашиваемость пыльцы раствором Люголя показала, что низкая фертильность пыльцы линий MS будет вызвана неполной деградацией крахмала во время созревания пыльцы ( Рис. 5 ). Было обнаружено, что семенная фертильность как при самоопылении, так и при обратном скрещивании была высокой во всех этих трех линиях ( Таблица 1 ). Линии МС показали высокий процент плодов с большим количеством семян в плодах, полученных самоопылением. Эти результаты подтвердили, что можно размножать линии МС путем самоопыления и производить чистые линии МС посредством культуры пыльников.
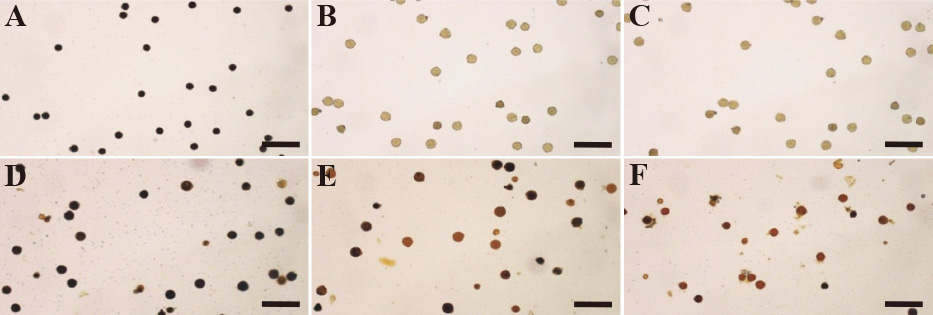
Рис. 4
Стадзависимые различия в окрашиваемости пыльцы в растворе Люголя между S. melongena 'Uttara' (A – C) и линиями мужской стерильности с цитоплазмой S. violaceum (D – F). За два дня до прохождения (A и D), в день прохождения (B и E) и через два дня после прохождения (C и F). Шкала бар = 100 мкм.
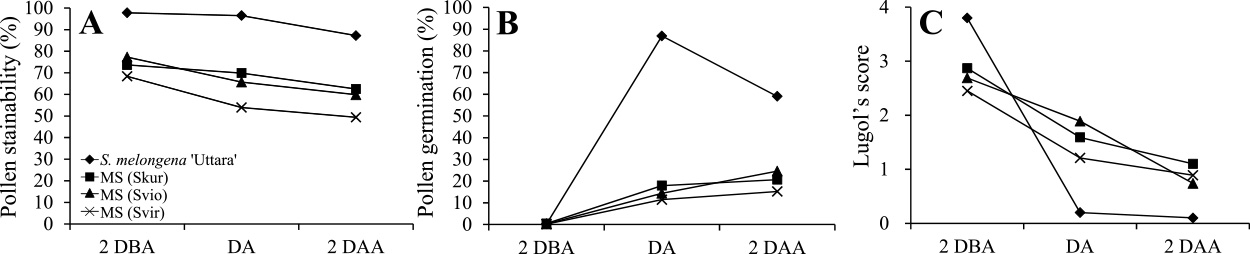
Рис. 5
Окрашиваемость пыльцы ацетокармином (A), прорастание пыльцы (B) и балл Люголя в трех системах мужской стерильности пыльников баклажана и у S. melongena 'Uttara', наблюдаемых за 2 дня до начала (2 дБА), в день приема ( DA) и через 2 дня после введения (2 DAA).
2) Характеристика пыльцевых систем мужского бесплодия
Характеристики пыльцы и фертильности семян линий MF исследовали в системах мужской стерильности пыльцы баклажана, разработанных с использованием цитоплазм S. aethiopicum Aculeatum Group, S. anguivi и S. grandifolium, чтобы выяснить возможности самоопыления и культура пыльников для развития гомозиготных реставраторов и линий МС ( Хан и др., 2013 ). Методы самоопыления и пылеобразования линий MF были применены, чтобы подтвердить полезность этих методов. Характер пыльцы линий MF оценивали аналогично, как показано в Рисунок 5 , Окрашиваемость пыльцы в ацетокармине и скорость прорастания пыльцы in vitro на всех трех стадиях созревания пыльцы были ниже в линиях MF, чем в баклажане «Уттара». В противоположность этому, было установлено, что степень окрашивания пыльцы в ацетокармине составляет около 50%, а всхожесть пыльцы in vitro составляет менее 20% в этих трех линиях MF. Окрашиваемость пыльцы в растворе Люголя показала, что низкая фертильность пыльцы линий MF будет вызвана неполным накоплением и деградацией крахмала во время созревания пыльцы. Было обнаружено, что плодородие семян обычно высоко в линиях МС пыльцевого типа ( Таблица 1 ). Линии MF показали хороший процент плодоношения и дали достаточное количество семян при самоопылении и обратном скрещивании. Двойные гаплоидные растения MS и MF были получены из культуры пыльников линий MF с цитоплазмами S. anguivi и S. grandifolium соответственно ( Таблица 3 ). Изучение фертильности пыльцы и семян линий MF показало, что гомозиготные реставраторы и линии MS могут быть получены путем самоопыления и пылеобразования линий MF систем мужской стерильности пыльцы без образования пыльцы баклажана.

Таблица 3
Частота образования эмбрионов из культивируемого пыльника линий мужской фертильности (MF) трех цитоплазматических систем мужской стерильности баклажана с цитоплазмами S. aethiopicum , S. anguivi и S. grandifolium .
3. Восстановление фертильности в системах CMS баклажана по генам Rf систем друг друга
Системы CMS традиционно характеризовались генами Rf, необходимыми для преодоления CMS и обеспечения потомства MF в цитоплазме MS ( МакВетти, 1997 ). Ген Rf до сих пор не обнаружен в других системах мужской стерильности баклажанов, разработанных с использованием цитоплазм S. kurzii , S. violaceum и S. virginianum . Однако в системах CMS были обнаружены гены Rf с цитоплазмами S. aethiopicum Aculeatum Group, S. anguivi и S. grandifolium ( Хан и др., 2014 ). Была оценена способность к восстановлению фертильности генами Rf систем друг друга в трех системах CMS типа пыльцы баклажана. Линия MS каждой системы CMS была опылена пыльцой линий MF других систем CMS, а также собственной системой CMS ( Рис. 6 ). Среди 9 перекрестных комбинаций 6 групп были получены путем скрещивания линий MS с линиями MF других систем CMS, и 3 группы были получены путем скрещивания линий MS с линиями MF их собственной системы CMS. Сегрегация растений MF и MS хорошо сочетается с соотношением 3: 1 почти во всех перекрестных комбинациях ( Таблица 4 ). Модели сегрегации показали, что два независимых доминантных гена Rf восстанавливают фертильность в каждой системе CMS. Окрашиваемость пыльцой и показатели прорастания пыльцы in vitro у потомства, полученного в различных кросс-комбинациях, варьировали в пределах 61,0–89,8% и 0,7–8,0% соответственно. Каждый ген Rf трех систем CMS был способен восстановить фертильность других систем CMS, а также свою собственную систему CMS. Более того, было обнаружено, что действия по восстановлению фертильности в этих трех видах CMS были схожими.
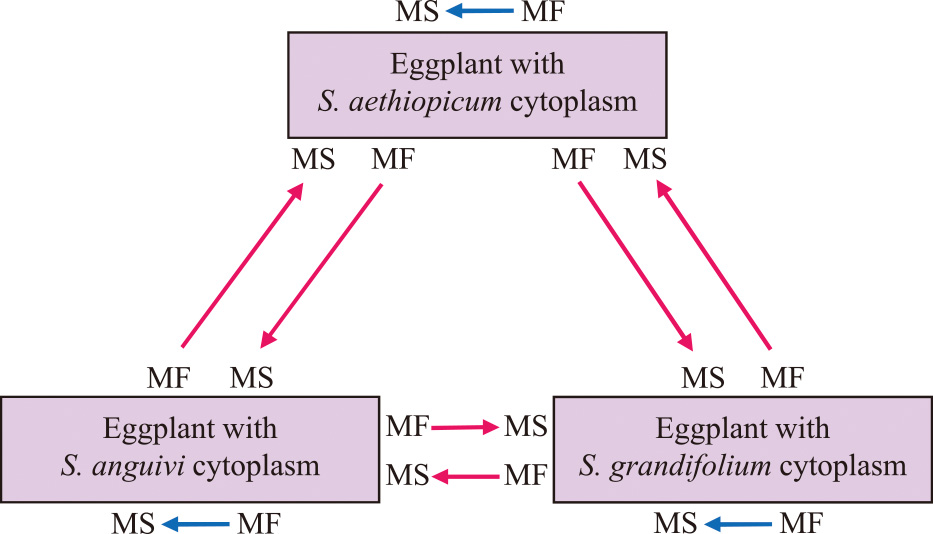
Рис. 6
Перекрестные комбинации между линиями мужской стерильности (MS) и мужской фертильности (MF) в трех системах мужской бесплодия пыльцы баклажана. Стрелки указывают направление опыления.
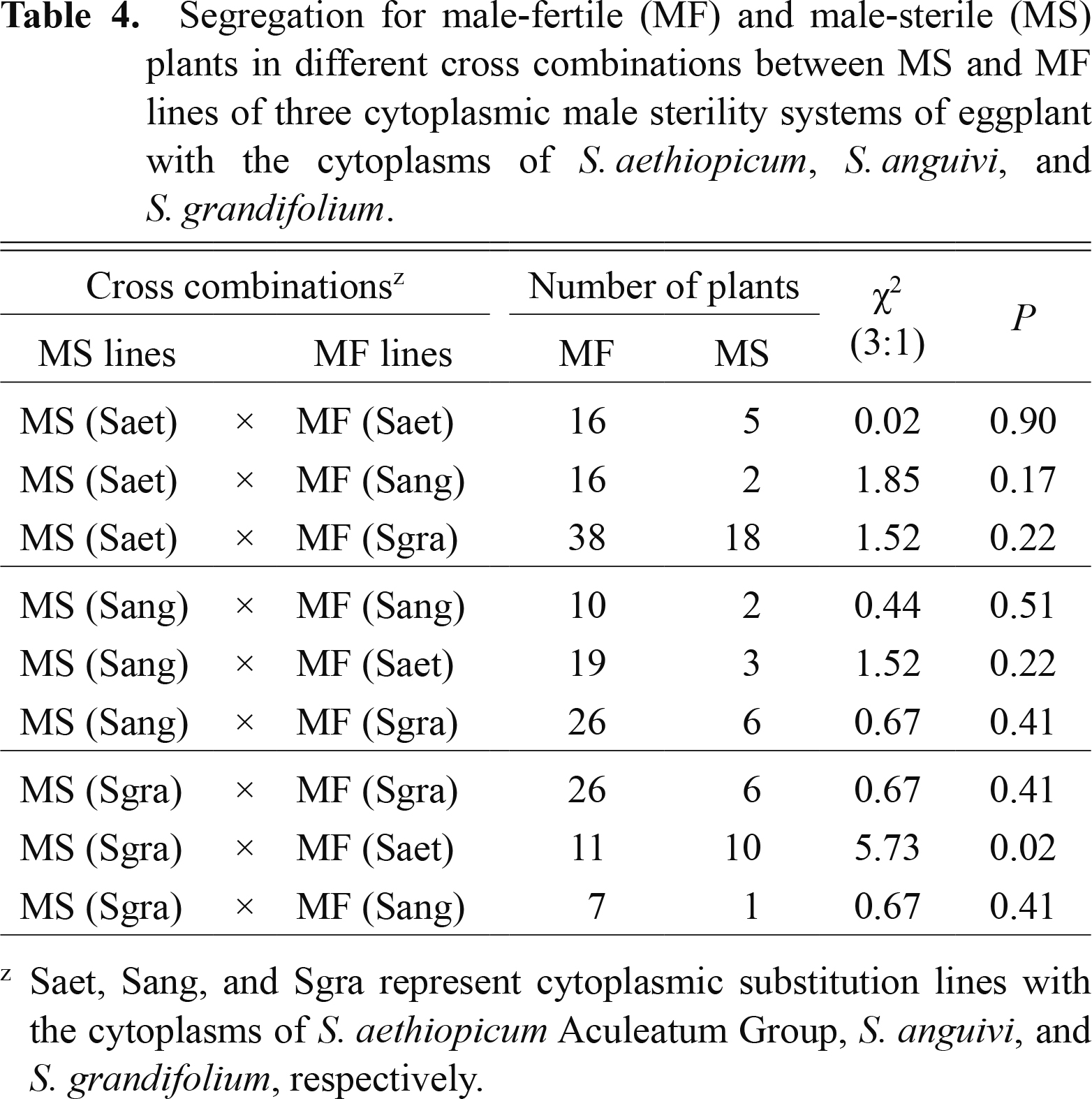
Таблица 4
Сегрегация для мужских фертильных (MF) и мужских стерильных (MS) растений в различных перекрестных комбинациях между линиями MS и MF трех цитоплазматических систем мужской стерильности баклажана с цитоплазмами S. aethiopicum , S. anguivi и S. grandifolium .
4. Разработка маркера СКАР, связанного с геном Rf
ДНК-маркеры, связанные с геном Rf , подвергали скринингу из трех систем CMS типа пыльцы баклажана с цитоплазмами S. aethiopicum Aculeatum Group, S. anguivi и S. grandifolium методом объемного сегрегации с использованием случайной амплифицированной полиморфной ДНК (RAPD). ) ( Хан и др., 2014 ). Аликвоты ДНК из 10 произвольно отобранных растений MF и MS из потомства обратного скрещивания наливали отдельно для каждой системы CMS. Объемные образцы ДНК были подвергнуты скринингу с использованием 220 видов праймеров Operon и 200 видов праймеров Common Bex от Operon Technologies (Alameda, CA, USA). Всего было использовано 420 праймеров, и более 65% праймеров давали значительное количество фрагментов RAPD как в массе MF, так и в массе MS. Только один праймер Operon, OPAB10, давал воспроизводимую полиморфную полосу 1,9 т.п.н. (OPAB101900) между объемами и был идентифицирован как тесно связанный с генами Rf растений MF. Кроме того, маркер RAPD OPAB101900 был преобразован в секвенированный охарактеризованный маркер амплифицированной области (SCAR), обозначенный как SCAB101900, путем нуклеотидного секвенирования ( Рис. 7 ). SCAB101900 может быть успешно использован для отбора с помощью маркера, и ожидается, что он поможет селекционерам отличать MF от растений MS до сброса пыльцы. Этот метод отбора может быть полезен для быстрого скрининга инбредных линий на ген Rf / rf без разработки и оценки потомства тесткросса. Этот маркер также может быть полезен для выделения генов Rf .
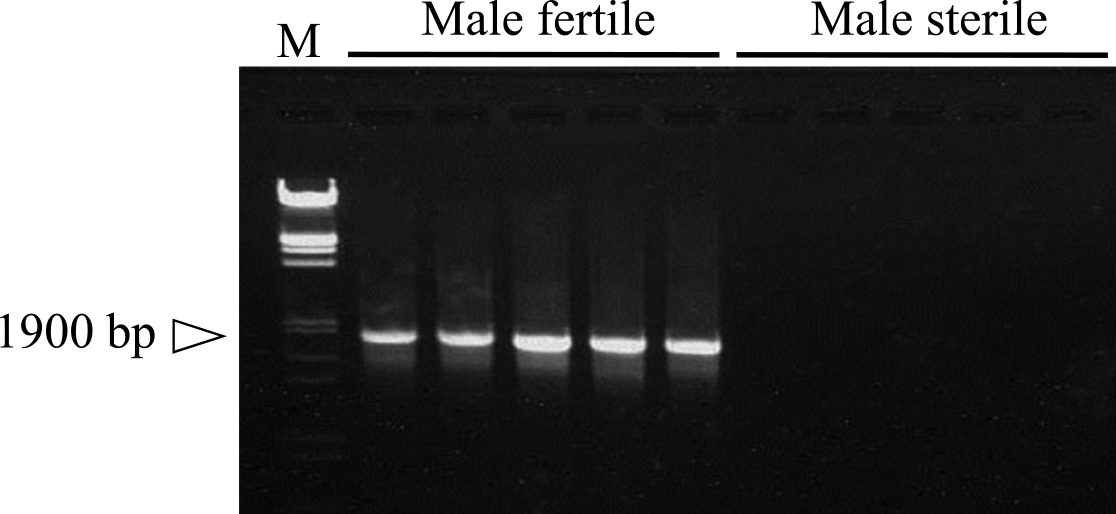
Рис. 7
Сегрегация маркера SCAR (SCAB101900) у мужчин с фертильной и мужской стерильностью у баклажанов с цитоплазмой S. anguivi . M является маркером двойного переваривания Eco / R, λ / Hin d III.
5. Заключение и перспектива
В этом обзоре обобщены исследования CMS на баклажанах с акцентом на развитие CMS, характеристику, восстановление фертильности и разработку маркера SCAR, связанного с генами Rf . Результаты исследований систем CMS в баклажанах могут дать нам возможность исследовать CMS и соответствующие гены Rf из диких ресурсов Solanum для программ разведения баклажанов ( Таблица 5 ). Выделение и изучение функции гена Rf может позволить лучше понять вовлеченные механизмы. Молекулярная основа CMS в аллоплазматических линиях баклажана была описана, где геномные структуры и паттерны транскрипции генов субъединицы митохондриальной АТФ-синтазы и субъединицы цитохромоксидазы были изучены для диких и культивируемых баклажанов ( Йошими и др., 2013 ). Диверсификация источников CMS необходима для использования в коммерческом производстве гибридных семян. Линии партенокарпических баклажанов были разработаны генной инженерией ( Acciarri et al., 2002 ; Донцелла и др., 2000 ; Ротино и др., 1997 ) и обычного скрещивания ( Кикучи и др., 2008 ; Сайто и др., 2009b ). Включение этих партенокарпических признаков в существующие системы CMS будет очень полезно для производства высококачественных плодов без косточек и для плодоношения гибридных сортов без обнаружения генов Rf или применения обработки фитогормонами. Наконец, необходимо расширить исследования механизмов, участвующих в восстановлении мужской стерильности и фертильности, чтобы улучшить наше понимание ядерных цитоплазматических взаимодействий и достичь лучшего использования систем CMS / Rf для гибридного размножения в баклажанах. Дальнейшие усилия должны быть предприняты, чтобы выяснить природу и роль кодируемых ядром белков в экспрессии важных митохондриальных генов. Со всеми этими ресурсами будущее исследований по разведению баклажанов обещает быть исключительно плодотворным.

Таблица 5
Резюме цитоплазматических систем мужской стерильности баклажана, найденных в нашем исследовании.
Литература цитируется
- Acciarri, Н., Ф. Restaino, Г. Вителли, Д. Перроне, М. Зоттини, Т. Пандольфини, А. Спена и Г. Л. Ротино. 2002. Генетически модифицированные партенокарпические баклажаны: повышение продуктивности плодов при выращивании в теплицах и в открытом грунте. BMC Biotechnol. 2 : 1–7.
- Бентолила С., Альфонсо А. А. и Хансон М. Р. 2002. Ген, содержащий пентатрикопептидный повтор, восстанавливает фертильность цитоплазматических мужских стерильных растений. Proc. Natl. Акад. Sci. США 99 : 10887–10892.
- Chauhan, SVS, 1984. Исследования по генной мужской стерильности Solanum melongena L. Indian J. Genet. 44 : 367–371.
- Донзелла Г., А. Спена и Г. Л. Ротино. 2000. Трансгенные партенокарпические баклажаны: превосходная зародышевая плазма для увеличения зимней продуктивности. Mol. Порода. 6 : 79–86.
- Клык М., Р. Мао и В. Се. 1985. Разведение цитоплазматически наследуемых мужских стерильных линий яичного растения ( Solanum melongena L.). Acta Hortic. Грех. 12 : 261–266 (на китайском с аннотацией на английском языке).
- Gotoh, K. 1952. Исследования сочетания способности на сортах баклажанов. Япония. J. Порода. 1 : 196.
- Хансон М.Р. и С. Бентолила. 2004. Взаимодействие митохондриальных и ядерных генов, которые влияют на развитие мужского гаметофита. Plant Cell 16 : 154–169.
- Хаснуннахар, М., MMR Хан и С. Иссики. 2012a. Наследование восстановления мужской фертильности системы мужской стерильности в баклажанах с цитоплазмой S. grandifolium . Aust. J. Crop Sci. 6 : 475–479.
- Хаснуннахар, М., MMR Хан и С. Иссики. 2012b. Пыльца и плодородие семян трех функциональных мужских стерильных линий баклажана с дикой цитоплазмой Solanum . Sci. Hortic. 139 : 58–61.
- Isshiki, S. и N. Kawajiri. 2002. Эффект цитоплазмы Solanum violaceum Ort. на плодовитость баклажана ( S. melongena L.). Sci. Hortic. 93 : 9–18.
- Jasmin, JJ 1954. Мужское бесплодие в Solanum melongena L .: предварительный отчет о функциональном типе мужского бесплодия у баклажанов. Proc. Amer. Soc. Хорт. Sci. 63 : 443.
- Какидзаки, Ю. 1930. Разведение скрещенных баклажанов в Японии. Дж. Херед. 21 : 253–258.
- Какидзаки, Y. 1931. Гибридная энергия в баклажанах и ее практическое использование. Генетика 16 : 1–25.
- Хан, MMR и С. Иссики. 2008. Разработка мужского стерильного баклажана с использованием цитоплазмы Solanum virginianum и бипарентной передачи ДНК хлоропласта при обратном скрещивании. Sci. Hortic. 117 : 316–320.
- Хан, MMR и С. Иссики. 2009. Функциональное мужское бесплодие, выраженное в баклажане ( Solanum melongena L.), содержащем цитоплазму S. kurzii Brace & Prain. Дж. Хорт. Sci. Biotech. 84 : 92–96.
- Хан, MMR и С. Иссики. 2010. Разработка линии мужской стерильности баклажанов с использованием цитоплазмы Solanum aethiopicum L. Aculeatum Group. J. Япония. Soc. Хорт. Sci. 79 : 348–353.
- Хан, MMR и С. Иссики. 2011. Разработка цитоплазматической линии мужской стерильности баклажана ( Solanum melongena L.) с цитоплазмой Solanum anguivi . Порода растений. 130 : 256–260.
- Хан, М.М.Р., М. Хаснуннахар, М. Ивайоши и С. Иссики. 2013. Пыльца и плодородие семян мужских фертильных линий, имеющих ген восстановления фертильности в трех системах CMS баклажана. Sci. Hortic. 157 : 39–44.
- Хан, М.М.Р., М. Хаснуннахар, М. Ивайоши и С. Иссики. 2014. Восстановление фертильности в трех системах CMS баклажана с помощью генов Rf систем друг друга и их маркера SCAR. Sci. Hortic. 172 : 149–154.
- Кикучи, К., И. Хонда, С. Мацуо, М. Фукуда и Т. Сайто. 2008. Устойчивость плодового набора вновь отобранных партенокарпических линий баклажанов. Sci. Hortic. 115 : 111–116.
- McVetty, PBE 1997. Цитоплазматическая мужская стерильность. п. 155-182. В кн .: К.Р. Шиванна и В.К. Соухни (ред.). Пыльцевая биотехнология для растениеводства и улучшения. Издательство Кембриджского университета, Кембридж, Великобритания.
- Нагаи, К. и С. Кида. 1926. Опыты по гибридизации различных штаммов Solanum melongena . Япония. J. Genet. 4 : 10–30 (на японском).
- Nuttall, VW 1963. Наследование и возможная полезность функциональной мужской стерильности у Solanum melongena L. Can. J. Genet. Cytol. 5 : 197–199.
- Phatak, SC и CA Jaworski. 1989. UGA 1-MS мужская стерильная зародышевая плазма баклажана. HortScience 24 : 1050.
- Phatak, SC, J. Liu, CA Jaworski и AF Sultanbawa. 1991. Функциональное мужское бесплодие в баклажане: наследование и связь с геном пурпурного цвета. Дж. Херед. 82 : 81–83.
- Ротино Г.Л., Э. Перри, М. Зоттини, Х. Соммер и А. Спена. 1997. Генная инженерия партенокарпических растений. Биотехнология природы. 15 : 1398–1401.
- Сайто, Т., Х. Мацунага, А. Сайто, Н. Хамато, Т. Кога, Т. Судзуки и Т. Йошида. 2009a. Новый источник цитоплазматической мужской стерильности и гена восстановления фертильности в линиях баклажанов ( Solanum melongena L.). J. Япония. Soc. Хорт. Sci. 78 : 425–430.
- Сайто, Т., Йошида, С. Монма, Х. Мацунага, Т. Сато, А. Сайто и Т. Ямада. 2009b. Разработка партенокарпического сорта баклажана 'Anominori'. JARQ 43 : 123–127.
- Самбандум, CN 1962. Гетерозис в баклажане ( Solanum melongena Linn.) Перспективы и проблемы в коммерческом производстве гибридных семян. Econ. Bot. 16 : 71–76.
- Schnable, PS и RP Wise. 1998. Молекулярные основы цитоплазматического мужского бесплодия. Trends Plant Sci. 3 : 175–180.
- Йошими, М., Ю. Китамура, С. Иссики, Т. Сайто, К. Ясумото, Т. Терачи и Х. Ямагиши. 2013. Вариации в структуре и транскрипции митохондриальных генов atp и cox у диких видов Solanum, которые вызывают мужскую стерильность в баклажане ( S. melongena ). Теор. Appl. Жене. 126 : 1851–1859.
Похожие
Домашние методы для проблем с простатойандропауза Мужская зона Профилактика мужчины В зависимости от пола, с возрастом мы больше подвержены различным заболеваниям. В случае мужчин, серьезный риск для здоровья, несомненно, является проблемой с простатой.